En el verano de 1965 el joven estudiante de biología Stuart Kauffman esperaba impaciente frente al centro de cálculo de la Facultad de Medicina de la Universidad de California. Por entonces los ordenadores eran herramientas esotéricas, sólo al alcance de unos pocos privilegiados. Por eso el joven había tenido que pagar de su bolsillo el tiempo de cómputo necesario para ejecutar una red booleana formada por cien elementos, con dos entradas aleatorias en cada uno. Con la arrogancia de la juventud, Kauffman se proponía poner de evidencia las debilidades de la teoría de la evolución darwinista. Y, contra todo pronóstico, acertó.
Todo empezó a raíz de un curso que había realizado un año antes en Berkeley sobre biología general, en el que se trataban algunos de los problemas de la embriología. Kauffman se había sentido fascinado por la forma en que los 100.000 genes que entonces se pensaba que componían el genoma humano se podían combinar para originar los 250 tipos celulares que constituyen la anatomía humana. El número de estados potenciales de 100.000 genes, con dos opciones por gen (activado y no activado), es de 2100000 ≈ 1030000. Es decir, existe un uno seguido de 30.000 ceros de posibles maneras de combinar 100.000 elementos que pueden encontrarse en dos estados. Ahora bien, cada tipo celular es el resultado de una combinación particular de genes ¿Cómo es posible, se preguntaba Kauffman, que la evolución por sí sola seleccione de tan alto número de estados únicamente los 250 necesarios para la supervivencia de la especie humana? La opinión mayoritaria sostenía que tales configuraciones correspondían a aquellas que habían sido seleccionadas por el mecanismo de mutación-selección. Kauffman, sin embargo, discrepaba.
Azar creadorPara responder a su pregunta Kauffman supuso que los genes se disponían como una red en la que cada gen se activaba o no en función de las entradas que recibía del resto de los genes. En esto coincidía con los estudios de genética molecular, que han demostrado que en el genoma de cualquier ser vivo hay una densa red de interacciones entre los genes. Lo revolucionario de su planteamiento consistía en asumir que este conjunto de interacciones se podía dar al azar y que, de esta forma, podía llegarse a algún tipo de comportamiento emergente. Esta atrevida hipótesis era la que quería confirmar con su simulación en aquel verano.
El diseño de este tipo de redes se encuadra dentro de lo que se conoce como “redes booleanas”, en las que cada nodo de la red se comporta como una función de lógica binaria. Supongamos que la activación de cada gen depende del efecto de otros dos. Si un gen requiere para activarse el ser estimulado por los otros dos simultáneamente, está simulando la función AND y si requiere el estímulo de sólo uno de los otros dos o de ambos simula la función OR. De esta manera se puede definir una red asignando a cada gen una función u otra. Para entender su funcionamiento veamos un caso muy sencillo: supongamos una red formada por tres elementos, de los cuales el primero, al que llamaremos A, está definido por una función OR, mientras que los dos siguientes, B y C, constan de una función AND. Esto implica que el elemento A se activará cuando esté activo el B, el C o ambos, mientras que el B requiere para activarse el que estén activos el A y el C, y de forma análoga la activación del C requiere que estén activos el A y el B. De esta forma hemos diseñado una red de Kauffman formada por tres elementos y con dos entradas por elemento, como se muestra en el recuadro adjunto.
La célula como ciclo límite de estadosPara modelar una red de Kauffman tenemos que establecer el número de elementos de la red, las conexiones de cada elemento con los demás y la función booleana de cada elemento. Una vez definida la red se asigna un valor 0 o 1 a cada elemento, esto es, se crean las condiciones iniciales, y se deja que el sistema evolucione por sí solo. Al hacerlo así veremos cómo los diferentes elementos toman un valor u otro en función de la arquitectura de la red. Al cabo de un tiempo de cómputo comprobaremos cómo algunos puntos se comportan como atractores puntuales y su estado no cambia a partir de entonces, mientras que otro conjunto de puntos oscila de forma regular creando lo que en dinámica se conoce como ciclo límite. De acuerdo con la hipótesis de Kauffman las células están configuradas por la acción de estos ciclos límite, es decir, cada célula es el resultado de la activación cíclica de un conjunto de genes.
Trabajando con diferentes redes de dos conexiones, Kauffman se dedicó a contar el número de ciclos límite de la red y obtuvo una regla empírica según la cual este número era aproximadamente igual a la raíz cuadrada del número de elementos. Lo cual quería decir que una red formada por 100.000 elementos tendría unos 316 ciclos límite. Ahora bien, hasta hace poco se pensaba que el genoma humano estaba formado por unos 100.000 genes y el número de tipos celulares distintos es de 250, razonablemente semejante al número de ciclos límite.
Tal coincidencia hizo que Kauffman se plantease la posibilidad de que el genoma de los seres vivos se comportase como una red booleana, en el sentido de que cada gen o conjunto de genes pueden activar o inhibir a otros, mientras que las células serían el resultado de la acción conjunta de los genes que integran un ciclo límite. Actualmente sabemos que el número de genes es mucho menor del esperado, del orden de unos 40.000. Lejos de invalidar la teoría de Kauffman este nuevo dato la reafirma aún más, en la medida en que un número menor de genes implica una intensa modulación entre ellos, tal y como preveía su modelo. Espoleado por esta intuición, Kauffman se dedicó a investigar en la literatura científica la relación entre el número de genes y el de células en diferentes especies, y constató alborozado que esta relación se ajustaba aproximadamente a su ley de la raíz cuadrada.
Las implicaciones del modelo de Kauffman son demasiado demoledoras para la ciencia ortodoxa como para que se les conceda todavía demasiado crédito. De ser ciertas, las células que constituyen los ladrillos de cada ser vivo no serían el producto de la acción lenta y tenaz de la evolución biológica, sino consecuencia de la combinación al azar de los genes que integran su genoma. Se trata de una cuestión apasionante para la genética, pero ¿tendrá alguna relación con la ecología? La respuesta no llegó hasta una década más tarde.
Redes booleanas en ecologíaStuart Pimm es un ecólogo de la Universidad de Tennessee (Knoxville) que combina el trabajo de campo con el teórico, lo que es muy poco frecuente. Considera las redes tróficas como una propiedad emergente de los sistemas complejos. Para él, hay un equilibrio entre el número de especies de una comunidad y la fuerza de las interacciones entre ellas. Para demostrar el carácter anti intuitivo de las redes tróficas, Pimm pone el siguiente ejemplo: “Los ecólogos estudian con frecuencia los pares depredador-presa y se podría imaginar que si se eliminara el depredador de una comunidad la presa se beneficiaría, ¿no?” Esto mismo le preguntaba el escritor Roger Levin. “Bueno” –continúa Pimm– “imagina que el depredador de la presa A también come una segunda especie, la presa B. Imagina que A y B son competidores, que comen las mismas hojas, por ejemplo, o anidan en los mismos árboles, algo de ese estilo. De modo que si elimina el depredador, la especie A puede salir perdiendo porque sufrirá una competencia mayor de la especie B.”
Trabajando con su colaborador Mac Post, Pimm intentó a finales de los años setenta construir comunidades ecológicas en un modelo informático, en el que añadían las especies una tras otra, cada una con sus parámetros matemáticos elegidos al azar. Como era de esperar, comprobaron que algunas especies conseguían hacerse un hueco en la red trófica mientras que otras desaparecían. Pero obtuvieron además otros dos resultados interesantes: En primer lugar, que podían formar comunidades de hasta un máximo de doce individuos, lo que implicaba que las comunidades con pocas especies son más fáciles de invadir que aquellas más numerosas. En segundo lugar, que a igual número de especies era más fácil invadir una comunidad joven que una madura, lo que de alguna manera sugiere que las comunidades tienden a formar una red invisible que repele a las especies invasoras. Ambas conclusiones resultan bastante intrigantes, puesto que en el modelo no había ninguna limitación de materia o energía que restringiera el número de especies.
El enigma Humpty DumptyPor aquella época, el ecólogo Jim Drake, colega de Stuart en la Universidad de Knoxville, trabajaba en un problema semejante. Su programa empezaba con 125 especies de plantas, herbívoros y carnívoros, y dejaba que el sistema evolucionara libremente, seleccionando al azar las especies que trataban de entran en la comunidad que se iba formando. Al final, siempre llegaba a una comunidad muy persistente con unas quince especies. Lo sorprendente era que si repetía el experimento con el mismo grupo de especies inicial obtenía otra comunidad persistente, pero diferente de la anterior. Aún más desconcertante era el hecho de que si intentaba ensamblar una comunidad a partir de los quince individuos que habían sido seleccionados en una simulación previa, la comunidad no funcionaba. Y, además, muchas de las especies que antes habían tenido éxito se extinguían.
Cuando Drake mostró este resultado a Pimm lo bautizaron como el “efecto Humpty Dumpty” (en alusión al personaje de un célebre acertijo infantil inglés) y desde entonces se ha convertido en uno de los enigmas más sorprendentes de la ecología teórica. Fue por entonces cuando Stuart Kauffman fue a la Universidad de Knoxville a dar una charla sobre redes booleanas y relieves adaptativos rugosos. Pimm, que había acudido a la charla con un moderado interés, se dio cuenta de que las redes booleanas podían aplicarse igualmente a los modelos tróficos de ecología. Sencillamente, lo que para Kauffman eran interacciones entre genes, para él eran interacciones entre especies. Las redes booleanas demostraban que alguna de las transiciones necesarias para ensamblar una comunidad no podían ocurrir, lo que daba una pista sobre la solución al misterio con el que se enfrentaban los ecólogos de Knoxville. “Eché a correr por el pasillo y golpeé la puerta de Jim gritando: ¡He encontrado a Humpty Dumpty!”, recordó Pimm más tarde.
En cualquier caso, el optimismo de Pimm ha de ser matizado. Perece evidente que la solución a los problemas de las simulaciones de redes tróficas en ecología ha de partir de las redes de Kauffman, aunque hasta la fecha no se ha encontrado una respuesta plenamente convincente. Como afirma el propio Pimm, “a medida que nos familiarizamos con las complejidades reales del comportamiento, notamos oleadas cada vez más largas que surgen de lo profundo de la red trófica”.
RECUADRO
Esperamos tu respuesta a los Retos Eco-LógicosLos lectores que quieran recoger el guante y participar con su respuesta en la resolución de estos retos eco-lógicos, pueden ponerse en contacto con el autor en la siguiente dirección de contacto:
José Gabriel Segarra Berenguer • Instituto de Enseñanza Secundaria La Asunción de Nuestra Señora • c/ Huertos y Molinos, 40 • 03203 Elche / Elx • Alicante • Correo electrónico:
josegsegarra@yahoo.esTambién es posible sumarse al foro de debate abierto en la página web de Quercus: www.quercus.es
RECUADRO
Una red booleana con tres elementos A (or) B (and) C (and)Tabla del gen A
A B C 0 0 0 1 0 1
1 1 0 1 1 1Tabla del gen B
B A C 0 0 0 0 0 1
0 1 0 1 1 1Tabla del gen C
C A B 0 0 0 0 0 1
0 1 0 1 1 1Evolución de la red en función del estado de los tres genes
Tiempo T Tiempo T+1
A B C A B C 0 0 0 0 0 0 0 0 1 1 0 0
0 1 0 1 0 0 0 1 1 1 0 0 1 0 0 0 0 0
1 0 1 1 1 0 1 1 0 1 0 1 1 1 1 1 1 1BibliografíaLewin, R. (1995). Complejidad. El caos como generador del orden. Tusquets. Barcelona.
Kauffman, S. (2003). Investigaciones. Complejidad, autoorganización y nuevas leyes para una biología general. Tusquets. Barcelo













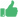 Si (
Si ( No(
No(





